
-
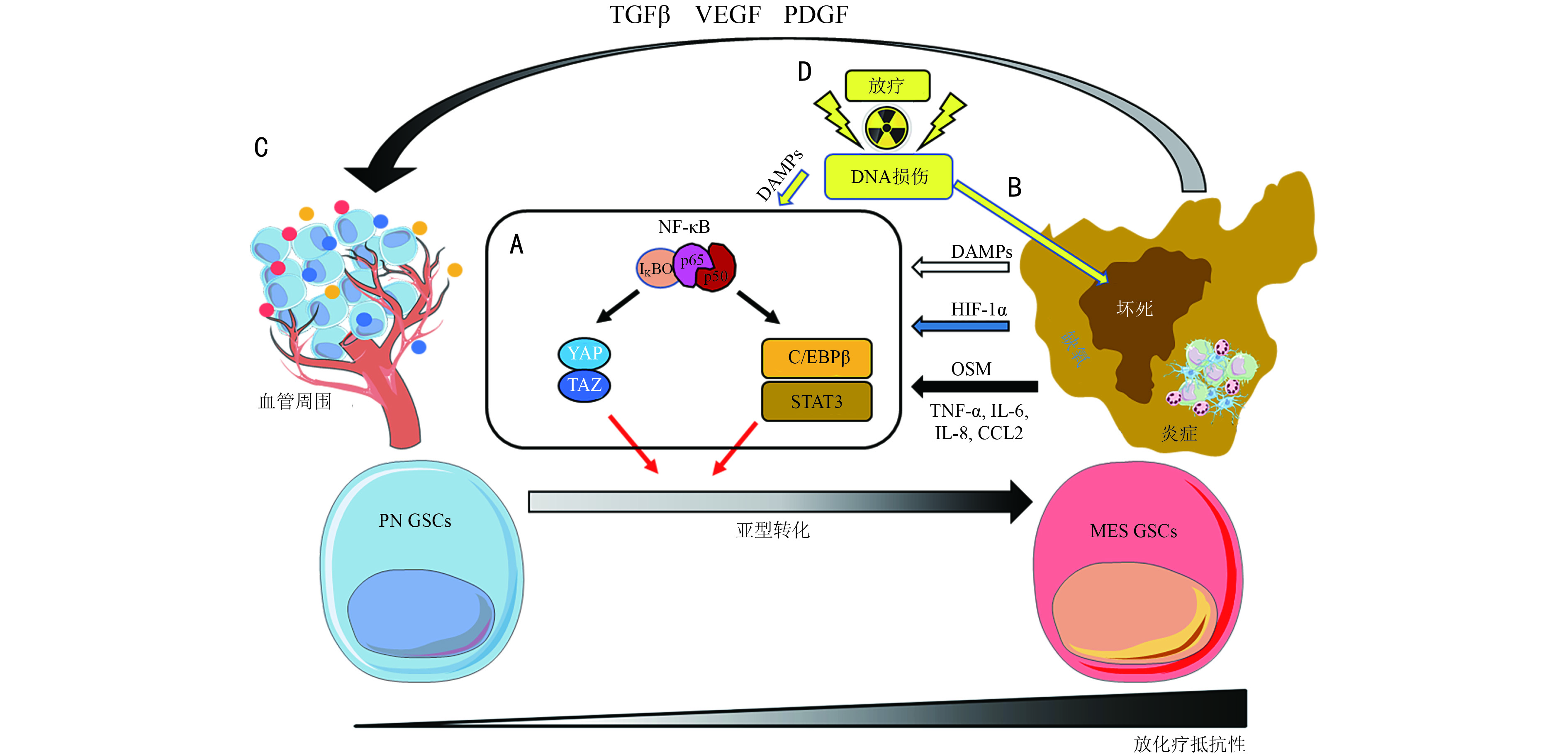
摘要: 胶质母细胞瘤(glioblastoma,GBM)是一种最常见恶性的原发性脑肿瘤,即使行标准化治疗后,患者中位总生存期仅12~16个月。GBM的治疗瓶颈主要来自于肿瘤的异质性。近年来,单细胞组学和细胞生物学研究的进展揭示了GBM中转录组亚型的混合和交互转化与其治疗效果密切相关。本文从GBM亚型分布特点,重点对恶性亚型(间充质亚型)形成及转化过程中的分子机制及环境因素进行综述,并讨论表型重塑相关机制,研究其在逆转胶质母细胞临床治疗抗性中的应用前景。Abstract: Glioblastoma is the most common and aggressive primary brain tumor, and patients with glioblastoma have a median overall survival of 12-16 months after standard therapy. The main complexity of glioblastoma therapy is tumor heterogeneity. Recent advances in single-cell and cell biology research indicate that the combination and cross-transition of transcriptome subtypes are related to therapeutic effects on glioblastoma. Thus, this study aimed to review the molecular mechanism and environmental factors related to the formation and transition of malignant glioblastoma subtypes (mesenchymal subtype) with respect to subtype distribution in the tumor microenvironment and integrate the mechanism into the clinical problem to discuss future glioblastoma therapeutic strategies.
-
Key words:
- glioblastoma /
- subtype transition /
- molecular mechanism /
- tumor microenvironment /
- clinical treatment
-
[1] Louis DN, Perry A, Wesseling P, et al. The 2021 WHO classification of tumors of the central nervous system: a summary[J]. Neuro Oncol, 2021, 23(8):1231-1251. [2] Teo WY, Sekar K, Seshachalam P, et al. Relevance of a TCGA-derived glioblastoma subtype gene-classifier among patient populations[J]. Sci Rep, 2019, 9(1):7442. [3] Guan XW, Vengoechea J, Zheng SY, et al. Molecular subtypes of glioblastoma are relevant to lower grade glioma[J]. PLoS One, 2014, 9(3):e91216. [4] Wang QH, Hu BL, Hu X, et al. Tumor evolution of glioma-intrinsic gene expression subtypes associates with immunological changes in the microenvironment[J]. Cancer Cell, 2018, 33(1):152. [5] van den Bent MJ, Gao Y, Kerkhof M, et al. Changes in the EGFR amplification and EGFRvⅢ expression between paired primary and recurrent glioblastomas[J]. Neuro Oncol, 2015, 17(7):935-941. [6] Wang JG, Cazzato E, Ladewig E, et al. Clonal evolution of glioblastoma under therapy[J]. Nat Genet, 2016, 48(7):768-776. [7] Hernández Martínez A, Madurga R, García-Romero N, et al. Unravelling glioblastoma heterogeneity by means of single-cell RNA sequencing[J]. Cancer Lett, 2022, 527:66-79. [8] Wang L, Babikir H, Müller S, et al. The phenotypes of proliferating glioblastoma cells reside on a single Axis of variation[J]. Cancer Discov, 2019, 9(12):1708-1719. [9] Patel AP, Tirosh I, Trombetta JJ, et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma[J]. Science, 2014, 344(6190):1396-1401. [10] Bhat KPL, Balasubramaniyan V, Vaillant B, et al. Mesenchymal differentiation mediated by NF-κB promotes radiation resistance in glioblastoma[J]. Cancer Cell, 2013, 24(3):331-346. [11] Fedele M, Cerchia L, Pegoraro S, et al. Proneural-mesenchymal transition: phenotypic plasticity to acquire multitherapy resistance in glioblastoma[J]. Int J Mol Sci, 2019, 20(11):E2746. [12] Carro MS, Lim WK, Alvarez MJ, et al. The transcriptional network for mesenchymal transformation of brain tumours[J]. Nature, 2010, 463(7279):318-325. [13] Bhat KP, Salazar KL, Balasubramaniyan V, et al. The transcriptional coactivator TAZ regulates mesenchymal differentiation in malignant glioma[J]. Genes Dev, 2011, 25(24):2594-2609. [14] Yamini B. NF-κB, mesenchymal differentiation and glioblastoma[J]. Cells, 2018, 7(9):E125. [15] Jin X, Kim LJY, Wu QL, et al. Targeting glioma stem cells through combined BMI1 and EZH2 inhibition[J]. Nat Med, 2017, 23(11):1352-1361. [16] Markwell SM, Ross JL, Olson CL, et al. Necrotic reshaping of the glioma microenvironment drives disease progression[J]. Acta Neuropathol, 2022, 143(3):291-310. [17] Guan F, Jiang WF, Bai Y, et al. Purinergic P2X7 receptor mediates the elimination of Trichinella spiralis by activating NF-κB/NLRP3/IL-1β pathway in macrophages[J]. Infect Immun, 2021, 59(5):e00683-e00620. [18] Zanoni M, Sarti AC, Zamagni A, et al. Irradiation causes senescence, ATP release, and P2X7 receptor isoform switch in glioblastoma[J]. Cell Death Dis, 2022, 13(1):80. [19] Uribe D, Niechi I, Rackov G, et al. Adapt to persist: glioblastoma microenvironment and epigenetic regulation on cell plasticity[J]. Biology (Basel), 2022, 11(2):313. [20] Wang ZL, Shi YP, Ying CT, et al. Hypoxia-induced PLOD1 overexpression contributes to the malignant phenotype of glioblastoma via NF-κB signaling[J]. Oncogene, 2021, 40(8):1458-1475. [21] Okuyama Y, Tanaka Y, Jiang JJ, et al. Bmi1 regulates IκBα degradation via association with the SCF complex[J]. J Immunol, 2018, 201(8):2264-2272. [22] Kim Y, Varn FS, Park SH, et al. Perspective of mesenchymal transformation in glioblastoma[J]. Acta Neuropathol Commun, 2021, 9(1):50. [23] Schmitt MJ, Company C, Dramaretska Y, et al. Phenotypic mapping of pathologic cross-talk between glioblastoma and innate immune cells by synthetic genetic tracing[J]. Cancer Discov, 2021, 11(3):754-777. [24] Hara T, Chanoch-Myers R, Mathewson ND, et al. Interactions between cancer cells and immune cells drive transitions to mesenchymal-like states in glioblastoma[J]. Cancer Cell, 2021, 39(6):779-792. [25] Niklasson M, Bergström T, Jarvius M, et al. Mesenchymal transition and increased therapy resistance of glioblastoma cells is related to astrocyte reactivity[J]. J Pathol, 2019, 249(3):295-307. [26] Pan YB, Wang SQ, Yang B, et al. Transcriptome analyses reveal molecular mechanisms underlying phenotypic differences among transcriptional subtypes of glioblastoma[J]. J Cell Mol Med, 2020, 24(7):3901-3916. [27] Brandes AA, Gil-Gil M, Saran F, et al. A randomized phase II trial (TAMIGA) evaluating the efficacy and safety of continuous bevacizumab through multiple lines of treatment for recurrent glioblastoma[J]. Oncologist, 2019, 24(4):521-528. [28] Chandra A, Jahangiri A, Chen W, et al. Clonal ZEB1-driven mesenchymal transition promotes targetable oncologic antiangiogenic therapy resistance[J]. Cancer Res, 2020, 80(7):1498-1511. [29] Minata M, Audia A, Shi JF, et al. Phenotypic plasticity of invasive edge glioma stem-like cells in response to ionizing radiation[J]. Cell Rep, 2019, 26(7):1893-1905. [30] Zhai K, Huang Z, Huang Q, et al. Pharmacological inhibition of BACE1 suppresses glioblastoma growth by stimulating macrophage phagocytosis of tumor cells[J]. Nat Cancer, 2021, 2(11):1136-1151. -
 下载:
下载:
 点击查看大图
点击查看大图
图(1)
计量
- 文章访问数: 190
- HTML全文浏览量: 49
- PDF下载量: 23
- 被引次数: 0
