
-
摘要:
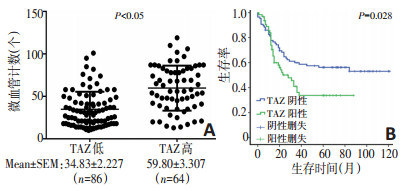
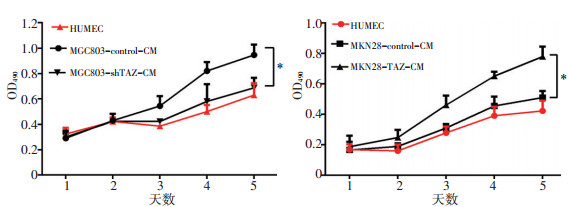
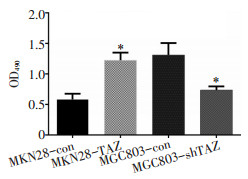
目的 探究Hippo通路关键效应分子TAZ在胃癌组织中的表达及其在胃癌血管生成中的作用。 方法 通过免疫组织化学法分析150例胃癌组织标本中TAZ和β-catenin的表达情况;将TAZ过表达质粒及干扰质粒通过慢病毒分别转染至胃癌细胞系MGC803和MKN28中,通过细胞功能实验检测内皮细胞成管、增殖及迁移能力;使用Western blot法检测转染后的胃癌细胞中TAZ及β-catenin的表达情况;采用酶联免疫吸附试验(ELISA)检测TAZ转染后血管内皮生长因子(vascular endothelial growth factor,VEGF)蛋白表达的变化。 结果 免疫组织化学法结果显示150例胃癌组织中,TAZ阳性表达64例(阳性率43%),主要定位于细胞核,其表达与肿瘤分级、TNM分期、转移及微血管密度(microvessel density,MVD)有关(P < 0.05)。此外,在TAZ阳性组中β-catenin阳性表达率为67.2%,明显高于TAZ阴性组,TAZ的表达与β-catenin呈正相关。在MKN28细胞系中上调TAZ的表达,与HUVEC细胞共培养后增强了内皮细胞增殖及管道形成能力,此外还通过促进β-catenin的表达,增强了内皮细胞的迁移能力;相反,在MGC803细胞系中下调TAZ的表达,与HUVEC共培养后减弱了内皮细胞增殖和管道形成能力,此外还通过降低β-catenin的表达,抑制了内皮细胞的迁移能力。 结论 胃癌细胞TAZ的高表达可能通过促进β-catenin和VEGF的表达,进而增强胃癌血管生成能力。 Abstract:Objective To determine the expression of TAZ and its role in angiogenesis in gastric carcinoma. Methods Immunohistochemical staining was performed to investigate the expression of TAZ and to determine whether a direct relationship exists between TAZ and β-catenin. Transfection with TAZ overexpression plasmid in MKN28 cells was conducted to induce exogenous expression of TAZ and a TAZ knockdown plasmid was transfected into MGC803 cells to reduce TAZ levels. The effects on endothelial cell formation, proliferation, and migration were determined by Matrigel three-dimensional culture, MTT proliferation assay and Transwell migration assay. In addition, the expression of TAZ and β-catenin in transfected gastric cancer cells was detected by Western blot. Results Immunohistochemistry showed that TAZ protein was expressed in 64 of 150 gastric cancer sample tissues (43%), TAZ was localized in the nucleus, and its expression was associated with tumor grade, TNM stage, metastasis, and microvessel density (MVD) (P < 0.05). In addition, the expression frequency of β-catenin in the TAZ positive group was 67.2%, which was significantly higher than that in the TAZ negative group, and the expression of TAZ was positively correlated with β-catenin. After transfection, TAZ overexpression increased the expression of β-catenin and enhanced HUVECs tube formation, proliferation, and migration. In the MGC803 cells transfected with the knockdown plasmid, β-catenin levels were decreased and HUVECs motility was inhibited. Conclusions TAZ may promote angiogenesis in gastric cancer by promoting β-catenin expression. -
Key words:
- gastric cancer /
- TAZ /
- β-catenin /
- angiogenesis /
- HUVEC cells
-

图 1 免疫组织化学法检测人胃癌组织中TAZ与β-catenin表达情况(×200)
A:TAZ阳性表达;B:TAZ阴性表达;C:β-catenin阳性表达;D:β-catenin阴性表达

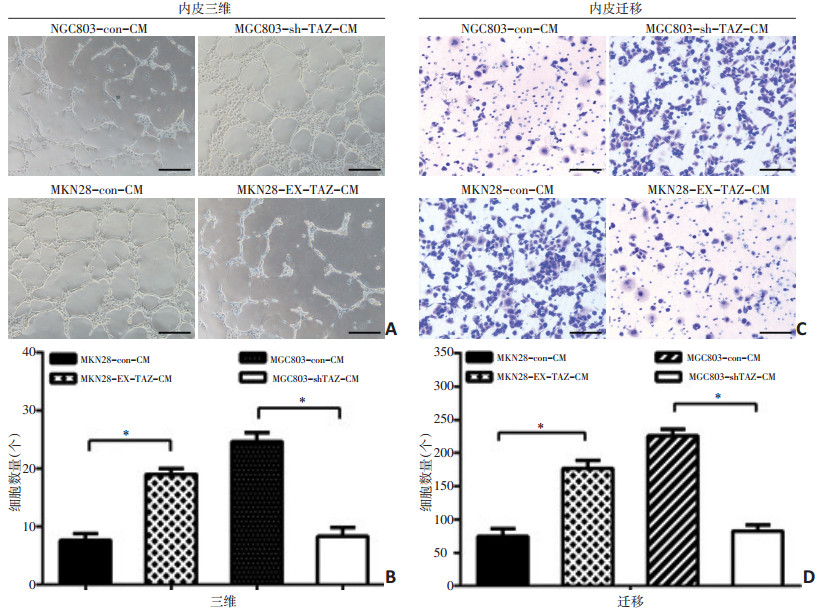
图 3 TAZ对内皮细胞管道形成能力及迁移能力的影响
A,B:过表达与降表达TAZ对内皮细胞管道能力的影响;C,D:过表达与降表达TAZ对内皮细胞迁移能力的影响;*:P<0.05

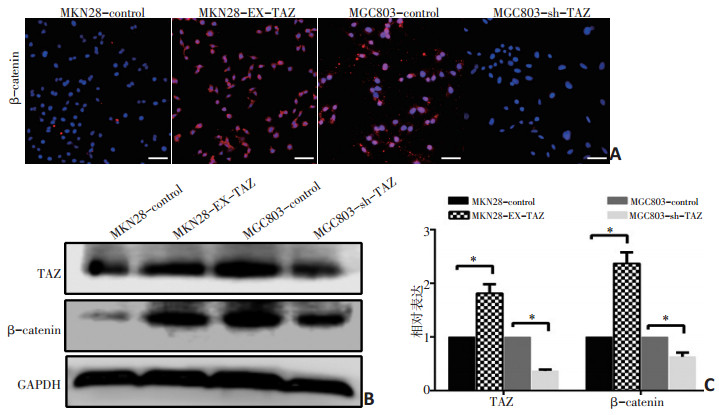
图 5 免疫荧光与Western blot检测TAZ与β-catenin的表达变化
A:免疫荧光检测TAZ转染后β-catenin的表达变化;B,C:Western blot法检测TAZ转染后TAZ与β-catenin的表达;*:P<0.05
表 1 TAZ与β-catenin的相关分析 例

表 2 TAZ的表达与胃癌临床病理特征的关系 例

-
[1] Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2018, 68(6):394-424. doi: 10.3322/caac.v68.6 [2] Katayama Y, Uchino J, Chihara Y, et al. Tumor Neovascularization and Developments in Therapeutics[J]. Cancers (Basel), 2019, 11(3):E316. doi: 10.3390/cancers11030316 [3] Schimmel L, Gordon E. The precise molecular signals that control endothelial cell-cell adhesion within the vessel wall[J]. Biochem Soc Trans, 2018, 46(6):1673-1680. doi: 10.1042/BST20180377 [4] Le Guelte A, Dwyer J, Gavard J. Jumping the barrier: VE-cadherin, VEGF and other angiogenic modifiers in cancer[J]. Biol Cell, 2011, 103(12): 593-605. doi: 10.1042/BC20110069 [5] Hu XY, Hou PF, Li TT, et al. The roles of Wnt/beta-catenin signaling pathway related lncRNAs in cancer[J]. Int J Biol Sci, 2018, (14):2003- 2011. [6] Ma R, Feng N, Yu X, et al. Promoter methylation of Wnt/beta-Catenin signal inhibitor TMEM88 is associated with unfavorable prognosis of non-small cell lung cancer[J]. Cancer Biol Med, 2017, 14(4):377-386. doi: 10.20892/j.issn.2095-3941.2017.0061 [7] Ahmed RA, Shebl AM, Habashy HO. Expression levels of beta-catenin and galectin-3 in meningioma and their effect on brain invasion and recurrence: a tissue microarray study[J]. Cancer Biol Med, 2017, 14(3): 319-326. doi: 10.20892/j.issn.2095-3941.2017.0024 [8] Clifford RL, Deacon K, Knox AJ. Novel regulation of vascular endothelial growth factor-A (VEGF-A) by transforming growth factor (beta)1: requirement for Smads, (beta)-CATENIN, AND GSK3(beta)[J]. J Biol Chem, 2008, 283(51):35337-35353. doi: 10.1074/jbc.M803342200 [9] Dilek FH, Topak N, Tokyol C, et al. Beta-Catenin and its relation to VEGF and cyclin D1 expression in pT3 rectosigmoid cancers[J]. Turk J Gastroenterol, 2010, 21(4):365-371. doi: 10.4318/tjg.2010 [10] Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the Roots of Cancer [J]. Cancer cell, 2016, 29(6):783-803. doi: 10.1016/j.ccell.2016.05.005 [11] Collak FK, Demir U, Ozkanli S, et al. Increased expression of YAP1 in prostate cancer correlates with extraprostatic extension[J]. Cancer Biol Med, 2017, 14(4):405-413. doi: 10.20892/j.issn.2095-3941.2017.0083 [12] Park JH, Shin JE, Park HW. The role of hippo pathway in cancer stem cell biology[J]. Mol Cells, 2018, 41(2):83-92. http://cn.bing.com/academic/profile?id=8116c6ce9e1add67fea33a328108c638&encoded=0&v=paper_preview&mkt=zh-cn [13] Wang X, Freire Valls A, Schermann G, et al. YAP/TAZ orchestrate VEGF signaling during developmental angiogenesis[J]. Dev Cell, 2017, 42(5): 462-478 e7. doi: 10.1016/j.devcel.2017.08.002 [14] Cho H, Kim J, Ahn JH, et al. YAP and TAZ negatively regulate prox1 during developmental and pathologic lymphangiogenesis[J]. Circ Res, 2019, 124(2):225-242. doi: 10.1161/CIRCRESAHA.118.313707 [15] Chan SW, Lim CJ, Guo K, et al. A role for TAZ in migration, invasion, and tumorigenesis of breast cancer cells[J]. Cancer Res, 2008, 68(8):2592- 2598. doi: 10.1158/0008-5472.CAN-07-2696 [16] Wang C, Gu C, Jeong KJ, et al. YAP/TAZ-mediated upregulation of gab2 leads to increased sensitivity to growth factor-induced activation of the PI3K pathway[J]. Cancer Res, 2017, 77(7):1637-1648. doi: 10.1158/0008-5472.CAN-15-3084 [17] Wang Y, Han Y, Guo Z, et al. Nuclear TAZ activity distinctly associates with subtypes of non-small cell lung cancer[J]. Biochem Biophys Res Commun, 2019, 509(3):828-832. doi: 10.1016/j.bbrc.2019.01.012 [18] Choi HJ, Zhang H, Park H, et al. Yes-associated protein regulates endothelial cell contact-mediated expression of angiopoietin-2[J]. Nat Commun, 2015, 6:6943. doi: 10.1038/ncomms7943 [19] Neto F, Klaus-Bergmann A, Ong YT, et al. YAP and TAZ regulate adherens junction dynamics and endothelial cell distribution during vascular development[J]. Elife, 2018, (7):e31037. https://www.ncbi.nlm.nih.gov/pubmed/29400648 [20] Hwang JH, Kim AR, Kim KM, et al. TAZ couples Hippo/Wnt signalling and insulin sensitivity through Irs1 expression[J]. Nat Commun, 2019, 10 (1):421. doi: 10.1038/s41467-019-08287-x [21] Imajo M, Miyatake K, Iimura A, et al. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/beta-catenin signalling[J]. EMBO J, 2012, 31(5):1109-1122. doi: 10.1038/emboj.2011.487 [22] Xing Y, Zhang Y, Jia L, et al. Lipopolysaccharide from Escherichia coli stimulates osteogenic differentiation of human periodontal ligament stem cells through Wnt/beta-catenin-induced TAZ elevation[J]. Mol Oral Microbiol, 2019, 34(1):1-13. [23] Karaman S, Leppanen VM, et al. Vascular endothelial growth factor signaling in development and disease[J]. Development, 2018, 145(14:): 1-8. http://d.old.wanfangdata.com.cn/OAPaper/oai_pubmedcentral.nih.gov_3286203 -
 下载:
下载:
 点击查看大图
点击查看大图

图(6) / 表(2)
计量
- 文章访问数: 68
- HTML全文浏览量: 2
- PDF下载量: 4
- 被引次数: 0
